![]()
The continental shelf is a relatively shallow region of the ocean (0-200 m deep) that is also biologically productive. The reason for this productivity is twofold – these shallow waters are often replete with sunlight and contain high concentrations of nutrients from multiple sources, including freshwater runoff from land and upwelling of deep waters from currents interacting with the shelf. The high nutrient and high light environment of the continental shelf is favorable for the growth of phytoplankton, which support all of the animals that rely on this primary productivity at the base of the food web. Shipping, fishing, tourism, and other human activities are all concentrated on the continental shelf due to both convenience and consistent biological productivity.
Despite the ecological, economic, and societal importance of continental shelves fostering diverse marine communities, key ocean dynamics relevant to life are still poorly understood. An example of a key process on shelf ecosystems is the change from a stratified or vertically structured water column to one that is well mixed. These changes can occur on a daily cycle (e.g., a strong wind event or hurricane completely mixes the water column) or across seasons (summer tends to have weaker winds that favor stratified environments). In the stratified scenario, temperature, salinity, dissolved oxygen and other oceanographic conditions change dramatically with depth, leading to formation of microhabitats that may favor certain types of organisms. In the well-mixed scenario, the temperature, salinity and dissolved oxygen will not change much as you move to deeper waters. Below are examples of temperature and chlorophyll-a profiles from different locations off the coast of Georgia USA (South Atlantic Bight). The color of the dot represents the buoyancy frequency (N2) at each depth, which is a measure of stratification intensity. Higher values mean stronger stratification and thus a more “layered” water column, which is also associated with peaks in chlorophyll-a (a proxy for phytoplankton abundance).
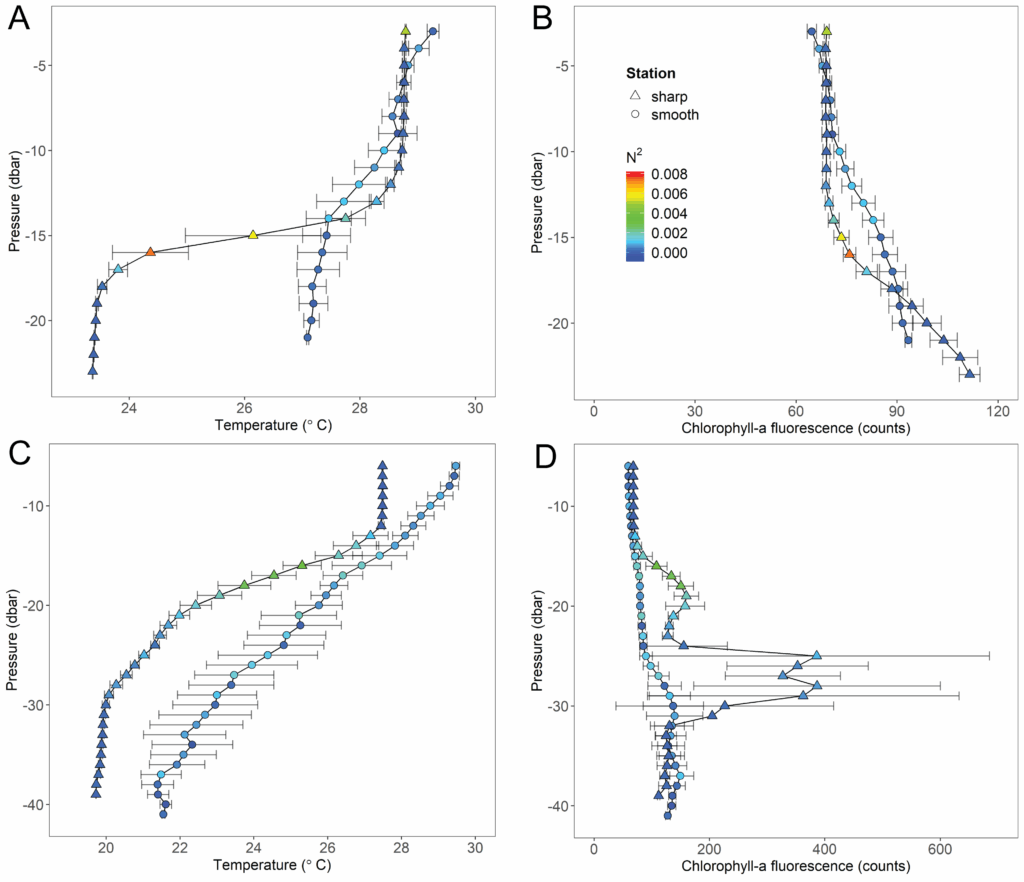
Vertical profiles from the South Atlantic Bight shelf during summer of 2021 collected with sensors on our modular Deep-focus Plankton Imager (mDPI, Greer et al. 2025). Note that sometimes the water is sharply stratified, and other times the thermocline gradually changes with depth, which indicates some degree of mixing (Aaron et al. 2025).

Similar reshuffling and community shifts occur in the ocean, yet our observations of these events (and the resulting changes throughout the food web) are limited by sampling capabilities. Conventional plankton nets are dragged through large portions of the water column (often 20 m or more) to measure abundances of zooplankton, which can “average out” oceanographic features and aggregations, such as those found in layers, that might be important for ocean animal ecology. Indeed, when high resolution sampling systems are used in shelf oceans (e.g., towed camera systems, such as the modular Deep-focus Plankton Imager (mDPI) shown below, Greer et al. 2025), they often show a water column that is highly structured (Greer et al. 2013; 2020; McManus et al. 2021). The patchiness or aggregations of organisms frequently generated by stratified conditions can also lead to increased efficiency of energy transfer to higher trophic levels (Greer and Woodson 2016). In other words, the productivity of our fisheries resources may also be linked to the formation of these layers and their dissipation. There is ample evidence that larger animals, such as fishes, dolphins, and whales, use layers and other ocean features as highly concentrated food resources (Benoit-Bird and McManus 2012; Fahlbusch et al. 2022). Thus, the oscillation between layers and well mixed conditions appears to influence the ecology of many ocean animals, but the consequences for the broader food web remain somewhat of a mystery.


Collecting in situ imaging data on the RV Savannah. The vehicle (above) is towed behind the ship and is connected to a winch and ultimately a shipboard computer. All data is transmitted from the vehicle to the computer via a fiber optic cable (yellow) where we can see the changes in the water column in real time. (And we try to make sure the vehicle doesn’t crash into the bottom of the ocean!)
In the NSF-funded DolLAYER project (Grant #2244690), we will be using a unique combination of technologies and sampling approaches to resolve the food web consequences for layered and well-mixed conditions. Specifically, we will be using in situ imaging to resolve the distributions of organisms and quantify how aggregated or dispersed they are in relation to layering and mixing. Next, we will use a new Multiple Opening and Closing Environmental Sampling System (MOCNESS) that can open and close nets at different depths (NSF Ocean Instrumentation Grant #2315030). We will use the MOCNESS and bottle samplers to collect organisms (from microbes to zooplankton) at specific depth ranges, informed by the imaging, to understand marine animal diets through both gut content and stable isotope analyses, and how this is related to the water column structure.
So what is the “Dol” in DolLAYER? This is an abbreviation of an animal called a “doliolid” that can be extremely abundant in continental shelf ecosystems. They look like little (~1 cm) gelatinous barrels, and they have a remarkable ability to reproduce and overwhelm the water column when conditions are favorable. (See an example .gif below collected with our benchtop imager – the darker organisms with antennae are copepods. Further down is a doliolid nurse, which is the larger life stage depicted in our DolLAYER logo.) We often refer to dense aggregations of doliolids as “blooms.” Recent work from our research team has suggested that these animals often feed on detritus (Walters et al. 2019; Frischer et al. 2021), which is a fancy word for dead and decaying material that is constantly sinking through the ocean (and yes, poop excreted by different animals is considered detritus). Are there any conditions that might favor dense aggregations of detritus? As you may have guessed, stratified water columns allow for sinking detritus to settle into layers, creating patches of detritus and hotspots for biological activity (Prairie et al. 2015).
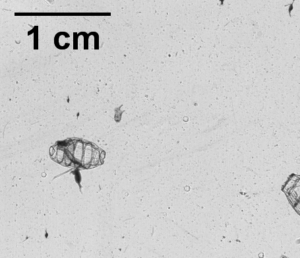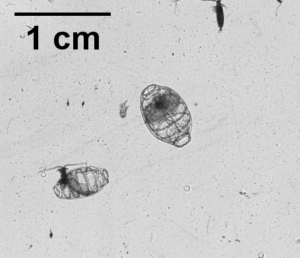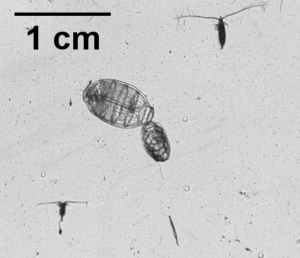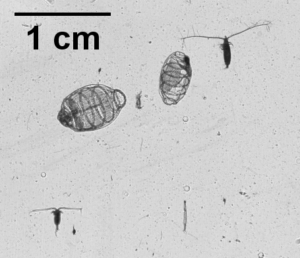
We hypothesize that these stratified conditions and layers may favor the proliferation of doliolids in continental shelf ecosystems, and this may lead to an overall gelatinous dominated food web because doliolid blooms might be a good food source for a variety of gelatinous predators (various medusae, ctenophores, and siphonophores – about which we know very little). Could this be a common occurrence in continental shelf oceans? Our combination of sampling methods should allow us to rigorously test this hypothesis.
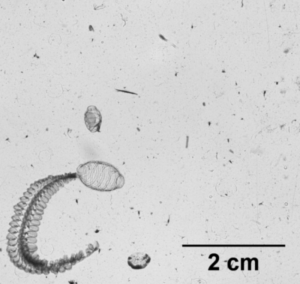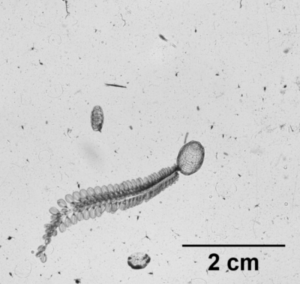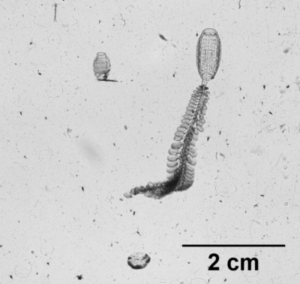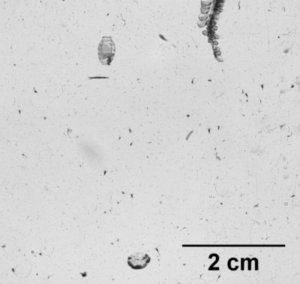
Furthermore, we will be conducting our sampling in the South Atlantic Bight and northern Gulf of Mexico – both are continental shelf ecosystems known to promote blooms of doliolids. The oceanographic mechanisms causing layer formation, however, are quite different between these two regions. The northern Gulf of Mexico receives large amounts of river discharge from a variety of sources (the Mississippi River is the most conspicuous), and this causes large salinity differences between the near surface (where the freshwater often comes in) and the deeper shelf waters that have higher salt content. The South Atlantic Bight, on the other hand, is less influenced by freshwater, so the main driver of layering is ocean temperature. In the South Atlantic Bight, these layers tend to form when cold, nutrient rich water is pushed onto the shelf, creating a strong temperature change between the surface water (heated by the sun) and these newly arrived colder waters. In the late fall and winter, both the northern Gulf of Mexico and the South Atlantic Bight tend to be well mixed.

Co-PI Dr. Laura Treible (Savannah State University) holds a cannonball jellyfish. Could these jellies feed on blooms of doliolids, or might they rely on layers in the water column?
Below are several links to help you get engaged with our project. Thank you for reading, and please reach out with any questions!
Publications and Resources for Teachers
The Doliolid Digest (research blog)
Zooplankton Identification Guide for the South Atlantic Bight
Cited References:
Aaron KD, Greer AT, Duffy PI, Treible LM, Frischer ME (2025) Stratification intensity structures zooplankton functional trait composition in a continental shelf system. ICES J Mar Sci 82(6): fsaf089
Benoit-Bird KJ and McManus MA (2012) Bottom-up regulation of a pelagic community through spatial aggregations. Biol Lett 8:813-816
Fahlbusch JA, Czapanskiy MF, Calambokidis J, Cade DE, Abrahms B, Hazen EL, Goldbogen JA (2022) Blue whales increase feeding rates at fine-scale ocean features. Proc. R. Soc. B 289: 20221180
Frischer ME, Lamboley LM, Walters TL, Brandes JA, Arneson E, Lacy LE, Lopez-Figueroa NB, Rodriguez-Santiago AE, Gibson DM (2021) Selective feeding and linkages to the microbial food web by the doliolid Dolioletta gegenbauri. Limnol Oceanogr 66:1993-2010
Greer AT, Cowen RK, Guigand CM, McManus MA, Sevadjian JC, Timmerman AHV (2013) Relationships between phytoplankton thin layers and the fine-scale vertical distributions of two trophic levels of zooplankton. J Plankton Res 35:939-956
Greer AT and Woodson CB (2016) Application of a predator – prey overlap metric to determine the impact of sub-grid scale feeding dynamics on ecosystem productivity. ICES J Mar Sci 73:1051-1061
Greer AT, Boyette AD, Cruz VJ, Cambazoglu MK, Dzwonkowski B, Chiaverano LM, Dykstra SL, Briseno-Avena C, Cowen RK, Wiggert JD (2020) Contrasting fine-scale distributional patterns of zooplankton driven by the formation of a diatom-dominated thin layer. Limnol Oceanogr 65:2236-2258
Greer AT, Duffy PI, Walles TJW, Cousin C, Treible LM, Aaron KD, Nejstgaard JC (2025) Modular shadowgraph imaging for zooplankton ecological studies in diverse field and mesocosm settings. Limnol Oceanogr: Methods 23(1): 67-86.
McManus MA, Greer AT, Timmerman AH, Sevadjian JC, Woodson CB, Cowen R, Fong DA, Monismith S, Cheriton OM (2021) Characterization of the biological, physical, and chemical properties of a toxic thin layer in a temperate marine system. Mar Ecol Prog Ser 678:17-35
Prairie JC, Ziervogel K, Camassa R, McLaughlin RM, White BL, Dewald C, Arnosti C (2015) Delayed settling of marine snow: Effects of density gradient and particle properties and implications for carbon cycling. Mar Chem 175:28-38
Walters TL, Lamboley LM, Lopez-Figueroa NB, Rodriguez-Santiago AE, Gibson DM, Frischer ME (2019) Diet and trophic interactions of a circumglobally significant gelatinous marine zooplankter, Dolioletta gegenbauri (Uljanin, 1884). Mol Ecol 28:176-189